4.1 METABOLISMO DE LIPIDOS
| Sitio: | Moodle ITST |
| Curso: | Bioquimica-M-P |
| Libro: | 4.1 METABOLISMO DE LIPIDOS |
| Impreso por: | Invitado |
| Fecha: | lunes, 28 de abril de 2025, 13:19 |
1. 3.1 METABOLISMO DE LIPIDOS
METABOLISMO DE LIPIDOS
Los principales lípidos de la ingesta son los triglicéridos, que están formados por 3 ácidos grasos esterificados al glicerol. Una vez que los ácidos grasos son liberados por acción de las lipasas, los ácidos grasos se degradan por una secuencia de reacciones que conforman la β-oxidación. Esta vía catabólica se realiza en la mitocondria de los organismos eucariontes y en el citosol de los procariontes. Los ácidos grasos que se degradan se encuentran en el citosol, en donde deben “activarse” por medio de una reacción de acilación para formar a la acil-CoA
La degradación de los ácidos grasos (β-oxidación). Generalidades. Productos de la vía y balance energético. Regulación
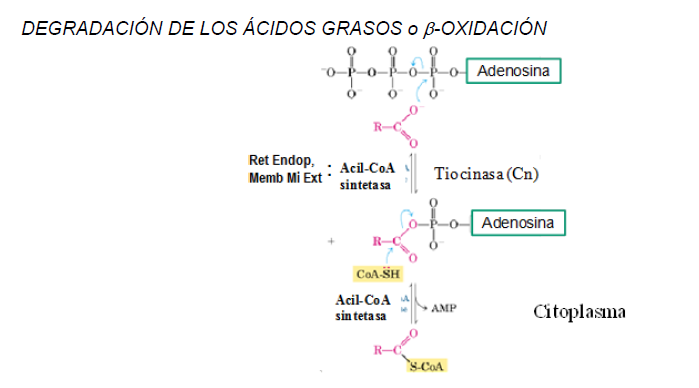
Activación del ácido graso. La acil-CoA sintetasa o tiocinasa, que se encuentra adosada al retículo endoplásmico o a la membrana externa mitocondrial, adiciona un AMP a partir de ATP a la molécula del ácido graso liberando un PPi. Después, la CoA reemplaza el enlace formado por el ácido graso con el AMP, se forma el acil-CoA y se libera AMP. Esta reacción está cercana al equilibrio, por lo que la hidrólisis de PPi por una pirofosfatasa inorgánica impulsa la reacción hacia la formación del producto.
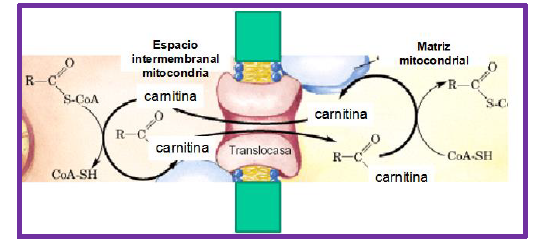
La entrada del acil-CoA a la mitocondria se realiza por medio de la carnitina. El enlace tioéster con la CoA es de una energía equivalente a la del enlace acil-carnitina, por lo que la carnitina palmitoil transferasa I reemplaza a la CoA por carnitina. El acil-carnitina se transporta hacia dentro de la mitocondria por un acarreador de acil-carnitina presente en la membrana mitocondrial. Ya adentro, la carnitina palmitoil transferasa II reconvierte el acil-carnitina en acil-CoA.
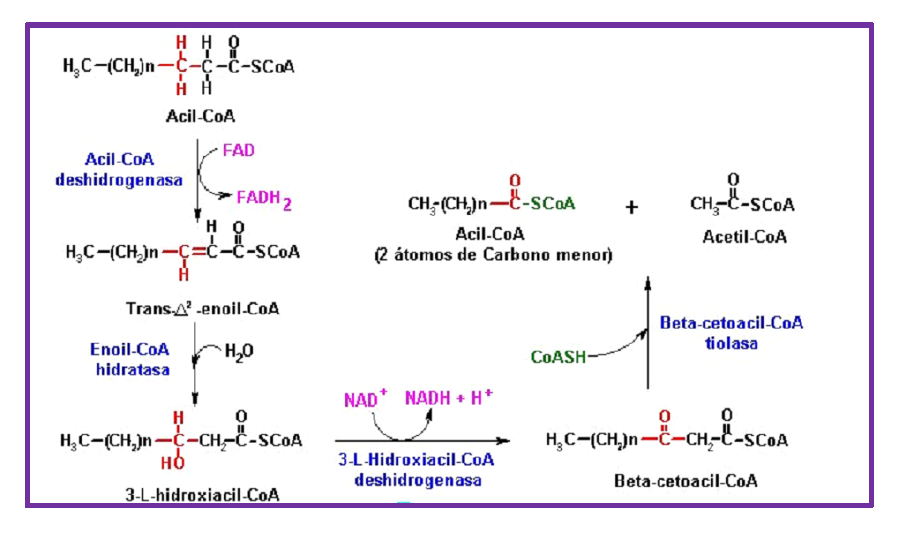
Reacciones de la β-oxidación.
La β-oxidación ocurre en cuatro reacciones:
1) La formación de un doble enlace trans entre los carbonos α y β del acil-CoA por una deshidrogenación catalizada por la acil-CoA deshidrogenasa. Esta reacción utiliza como cofactor el FAD+ que se convierte en FADH2. La enzima se reoxida por medio de la cadena transportadora de electrones, donde participa el complejo ETF (flavoproteína transferidora de electrones) y el complejo ETF:ubiquinona oxidoreductasa.
2) La hidratación del doble enlace por la enoil-CoA hidratasa para formar 3-L-hidroxiacil-CoA.
3) La deshidrogenación del 3-L-hidroxiacil-CoA por la 3-L-hidroxiacil-CoA deshidrogenasa con la formación de β-cetoacil-CoA. En esta reacción se genera NADH que también se reoxida por fosforilación oxidativa.
4) La ruptura del enlace Cα-Cβ por una tiolisis catalizada por la β-cetoacil tiolasa que genera un acetil-CoA y un acil-CoA con dos átomos de carbono menos.
Balance energético.
Cada vuelta de la β-oxidación genera un FADH2 y un NADH. Asimismo, el acetil-CoA generado puede entrar a Ciclo del ácido cítrico para generar 3 NADH, 1 FADH2 y 1GTP. Considerando que el FADH2 y el NADH generan, por oxidación en la fosforilación oxidativa 2 y 3 ATP´s, respectivamente:

Como puede verse, la oxidación completa de un ácido graso es altamente exergónica, por lo que genera gran cantidad de ATP. La oxidación del palmitoil-CoA (ácido graso de 16C) involucra 7 vueltas de β-oxidación generando 7 FADH2, 7 NADH y 8 acetil-CoA. La oxidación del acetil-CoA produce 8 GTP, 24 NADH y 8 FADH2. Restando los ATP requeridos para la formación del acil-CoA (en la primera reacción de activación del ácido graso), la oxidación de una molécula de palmitato genera 129 ATP.
La síntesis de ácidos grasos. Generalidades. Productos de la vía y balance energético. Regulación
Regulación hormonal de la oxidación del ácido graso
La oxidación de los ácidos grasos está regulada por la cantidad de ácidos grasos en sangre, que se liberan de los triacilgliceroles de tejido adiposo. Esta movilización se realiza por la lipasa de triacilglicerol dependiente de hormona. Esta enzima se regula por fosforilación-desforforilación dependiente de AMPc, que se controla hormonalmente. El glucagon y la epinefrina incrementan los niveles de AMPc, que activa a PKA, que fosforila y activa a la lipasa. Su actividad incrementa la concentración de los ácidos grasos en sangre, que son la señal para incrementar la degradación de lípidos en músculo e hígado. La insulina promueve la disminución del AMPc, por lo que promueve la desfosforilación de la lipasa. Asimismo, la insulina promueve la fosforilación independiente de la acetil-CoA carboxilasa, haciéndola más activa y estimulando la síntesis de los ácidos grasos.
Síntesis de los ácidos grasos
Al contrario de la β-oxidación, la síntesis de los ácidos grasos se lleva a cabo en el citosol y requiere la inversión de poder reductor biosintético en forma de NADPH. El precursor de la síntesis de ácidos grasos es el acetil-CoA, aunque el donador del C es el malonil-CoA.
Formación de malonil-CoA. La primera reacción involucra la carboxilación del acetil-CoA para formar un donador de grupos acetilo, el malonil-CoA.
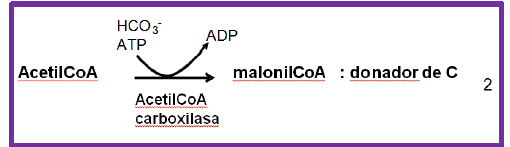
Esta reacción la cataliza la Acetil-CoA carboxilasa, que requiere la hidrólisis de un ATP y utiliza biotina como cofactor para la carboxilación y HCO3- como donador del carbono al acetil-CoA.
Esta reacción es la primera reacción reguladora de la síntesis de los ácidos grasos.
Reacciones subsecuentes. La síntesis de los ácidos grasos continúa con un conjunto de siete reacciones catalizadas por la ácido graso sintasa, que en animales es una enzima homodimérica multifuncional de 500 kDa. En E. coli, la síntesis se realiza por 7 enzimas diferentes, así como en las plantas, en donde las reacciones ocurren dentro del cloroplasto.
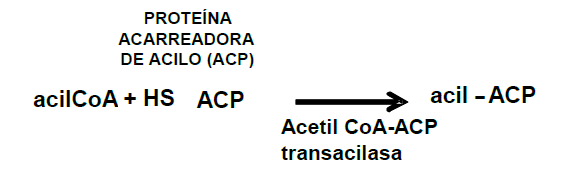
Para iniciar la síntesis del ácido graso se requiere que la cadena creciente esté esterificada a la Proteína Acarreadora de Acilo (ACP). Esta reacción es catalizada por la acetil-CoA-ACP transacilasa
La siguiente reacción la cataliza la β-cetoacil-ACP sintasa, que condensa un grupo acetil-ACP con uno de malonil-ACP (producido a partir de malonil-CoA por la malonil-CoA-ACP transacilasa). En esta reacción se libera una molécula de CO2, proveniente del malonil-ACP. El producto es un β-cetoacil-ACP (acetoacetil-ACP, en la primera vuelta de síntesis), éste se reduce por la β-cetoacil-ACP reductasa, que utiliza un NADPH como cofactor.
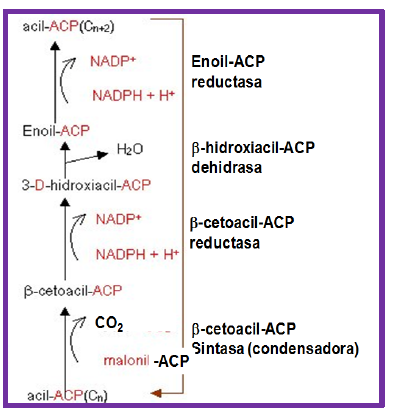
El 3-D-hidroxiacil-ACP generado es deshidratado por la β– hidrociacil-ACP dehidrasa para generar un doble enlace entre el Cα y Cβ, formando un enoil-ACP. Utilizando nuevamente NADPH, la enoil-ACP reductasa reduce el doble enlace para formar el acil-ACP con dos átomos de carbono más, acil-ACP(Cn+2). Éste vuelve a condensarse con otro acetil-ACP para que la cadena siga creciendo hasta formar palmitoil-ACP.
El palmitato (ácido graso de 16C) es el producto normal de la síntesis de los ácidos grasos, por lo que después de siete ciclos de síntesis, la palmitoil tioesterasa hidroliza el enlace tioéster con la ACP y libera palmitato. La reacción general de la síntesis del palmitato es:

La síntesis de ácidos grasos es energéticamente costosa.
El palmitato es el precursor de los ácidos grasos de cadena más larga y de ácidos grasos insaturados. Estas modificaciones son realizadas por las elongasas y las desaturasas, respectivamente.
Regulación del metabolismo de ácidos grasos.
La acetil-CoA carboxilasa es la primera enzima reguladora de la síntesis de los ácidos grasos. Su actividad se regula por polimerización. La forma polimérica es la más activa. El citrato estimula la formación del polímero por lo que se promueve la síntesis de los ácidos grasos. Mientras que el palmitoil-CoA promueve la despolimerización, por lo que se disminuye la síntesis. También está regulada a nivel hormonal por glucagon y epinefrina , que activan a PKA, que fosforila los protómeros de la acetil-CoA carboxilasa impidiendo la formación del polímero. Mientras que la insulina promueve la fosforilación en un sitio distinto, lo que promueve la polimerización.
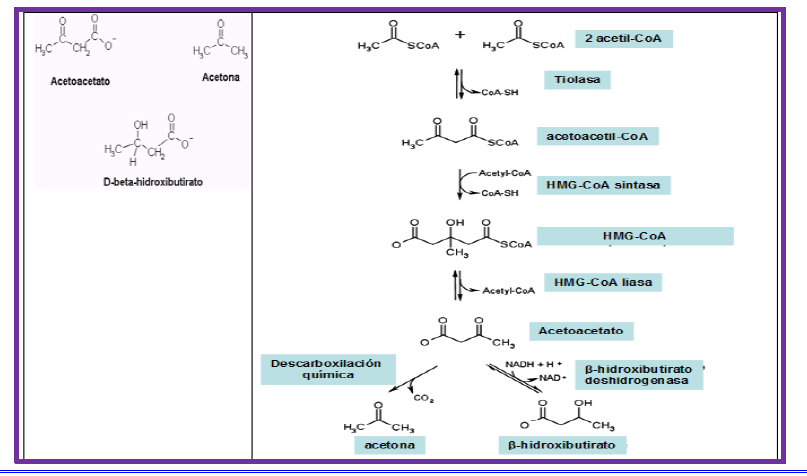
Síntesis de cuerpos cetónicos
Los cuerpos cetónicos son compuestos químicos producidos a partir de acetil-CoA proveniente de la β-oxidación. Este proceso se llama cetogénesis y se realiza en las mitocondrias del hígado, estos cuerpos cetónicos son una forma de almacenar acetil-CoA. La función de estos compuestos es suministrar energía a corazón, músculo esquelético y cerebro en ciertas situaciones excepcionales como ayuno muy prolongado, ya que generan acetil-CoA que puede oxidarse.
Los cuerpos cetónicos son el ácido acetoacético (acetoacetato) y el ácido betahidroxibutírico (β-hidroxibutirato); una parte del acetoacetato sufre descarboxilación no enzimática a acetona (una cantidad insignificante en condiciones normales)